








One of the most remarkable properties of the brain is its capacity to adapt at a functional and structural level in response to our environment and life experiences-it’s what allows us to learn, create memories, acquire skills, and adjust our behavior. This malleability of the brain is known as neuroplasticity or brain plasticity.
Until recently, it was believed that it was mostly at a functional level-through synaptic plasticity-that the adult brain could change. But the development and refinement of brain imaging technology has indicated that neuronal networks in the adult brain retain the ability to structurally remodel their connectivity-through synaptic remodeling or morphological neuronal adaptations, for example.
In this article, we take a look at the main mechanisms of functional and structural brain plasticity. The responsiveness of brain plasticity to the environment and life experiences is a topic we look into with more detail in our accompanying piece, Rewire Your Brain: Science-Backed Methods to Boost Neuroplasticity.
Neuroplasticity Acts from the Molecular to the Structural Level
As mentioned above, brain plasticity operates at two levels: functional and structural. Functional plasticity refers to the brain’s ability to adjust the patterns of responsiveness and activity of neurons, synapses, and neural circuits. It acts at the molecular level, mainly through modifications in presynaptic neurotransmitter release, postsynaptic receptor levels, intracellular signaling pathways, gene expression, and protein synthesis, which together influence the magnitude of synaptic transmission and neuronal firing. Structural plasticity refers to the brain’s ability to adjust neuronal morphology and structural aspects of neuronal connectivity. It acts at the cellular level through dynamic changes in cellular structures that have functional consequences.
Because of the type of changes they entail, functional plasticity is more pervasive and structural plasticity more limited. In fact, a high degree of structural invariability is an important characteristic of the mammalian central nervous system (CNS), as it ensures that neural circuits remain stable and that the specificity of neural connections is protected [1]. For that reason, neurons are largely unrenewable and neural connections are largely invariant, which is why the mammalian CNS has a very low capacity for regeneration and repair. But adaptability is as important as stability for the brain to function at full power. The secret is in a balance between stability and plasticity that allows us to grow with the environmental challenges that come our way.
An important attribute of the mechanisms of neuroplasticity is their responsiveness to our life experiences-what our senses capture from our environment and what we live through. That’s the essence of learning and memory: through long-lasting changes in neuronal communication and structure, our experiences get engraved into our brain and allow us to adapt our behavior [2].
Brain Plasticity Declines With Age
It’s common knowledge that children learn much faster than adolescents and adults. That’s because the brain is at its most plastic during developmental years, when neural circuits are still being formed and maturing. Children are able to absorb new information and develop new skills at an amazing rate because their brain is in a “critical period” of peak plastic capacity [3].
From adolescence onwards, plasticity starts to decline, particularly at the structural level, because the central nervous system starts to favor the stability and specificity of neural circuits as a way to retain the knowledge we’ve acquired [2]. But although the brain becomes less plastic as we age, a significant degree of plasticity, including structural plasticity, still persists throughout life.
Plasticity is a fundamental property of the brain. Despite declining with age, it is retained throughout life.
Mechanisms of Functional Plasticity
Functional plasticity acts at the molecular level through modifications in neurotransmitter signaling, receptor levels, ion channel properties, and gene expression, among others. Together, those adaptations influence the patterns of neural responsiveness and activity of neurons and neural circuits.
Synaptic strengthening
Functional synaptic plasticity consists in alterations in the signaling properties of synapses, most often in the form of changes in synaptic strength in response to patterns of neuronal activity, either by strengthening or weakening synaptic communication between two neurons. [See Figure 1 for a summary of a few basic concepts about neurons and synapses.]
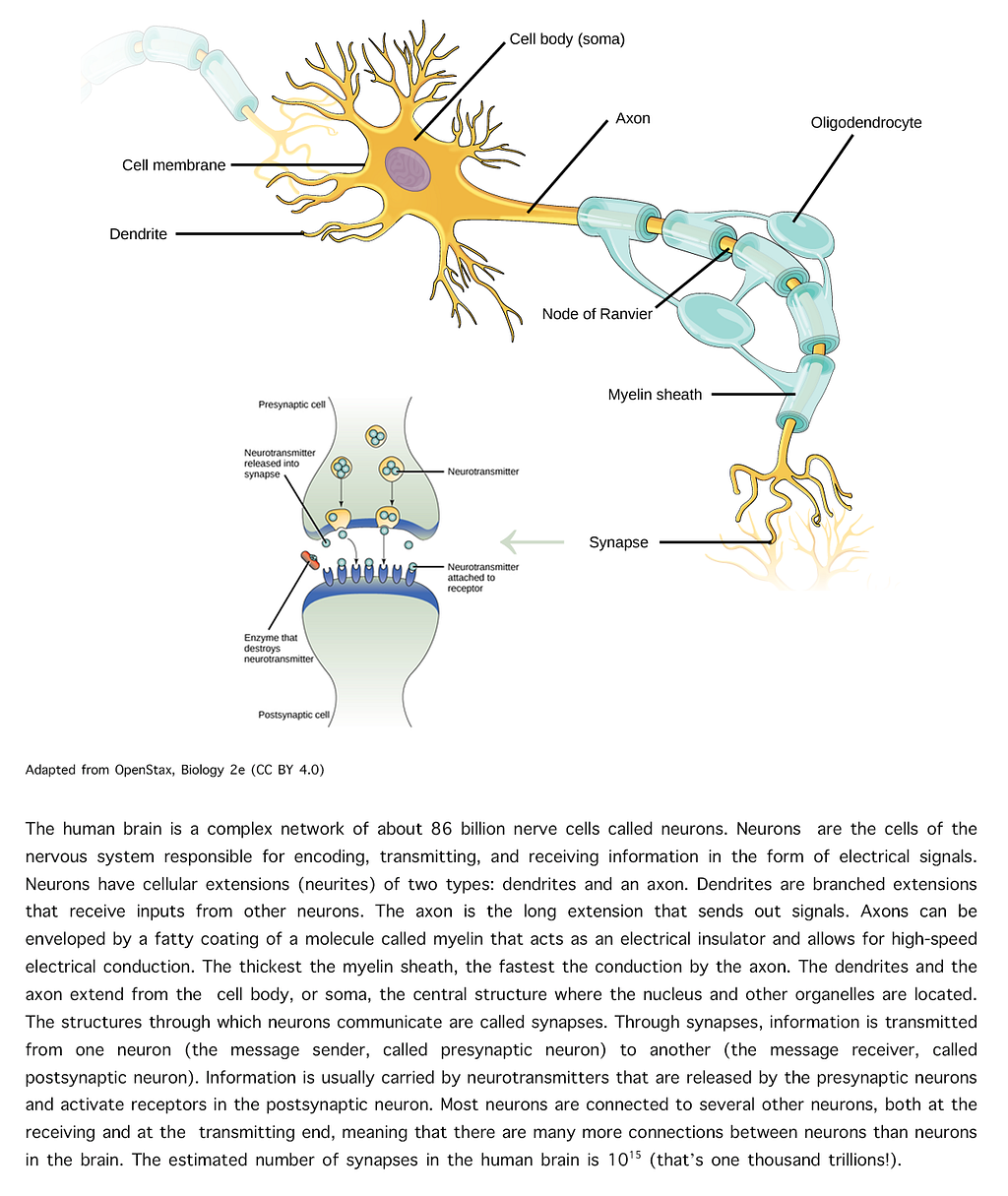
Figure 1 — Structure of neurons and synapses. Image adapted from OpenStax, Biology 2e (CC BY 4.0)
Synaptic strengthening means that, in response to increased activity, synapses become more efficient in transmitting information and eliciting a response in the postsynaptic neuron, thus strengthening the communication between neurons. The main form of synaptic strengthening is known as long-term potentiation (LTP). In this process, when two neurons fire in sequence, the responsiveness of the postsynaptic neuron is increased, making it more likely that it will fire the next time the presynaptic neuron fires. In an early phase, LTP involves the activation of signaling pathways and recruits existing molecules within neurons. As LTP becomes persistent, changes in gene expression take place that lead to the production of new proteins and the facilitation of neuronal communication [4,5]. Synaptic weakening, as in the case of long-term depression (LTD), is the opposite process: in response to decreased activity, synapses become less efficient and less able to elicit a response in the postsynaptic neuron, thus weakening communication between those two neurons.
LTP is one of the most relevant molecular mechanisms of learning and memory. Through LTP, relevant synaptic connections within a neuronal circuit activated by an experience are reinforced to become more efficient. This creates a “trace” of neuronal activity in the brain that acts a imprint of our experiences as it is integrated into our preexisting knowledge. The more an experience is repeated, the more permanent those synaptic changes become. That’s why you become gradually better at a skill with practice-you’re reinforcing the circuit of that memory.
[If you want to learn more about the mechanisms of LTP check out our article What is Glutamate? An Examination of the Functions, Pathways and Excitation of the Glutamate Neurotransmitter.Intrinsic Plasticity
Synaptic communication is the process of transmitting information from one neuron to another. That information may then be conveyed by the postsynaptic neuron in the form of an action potential. [See Figure 2 for a summary of a few basic concepts about action potential generation and transmission.]
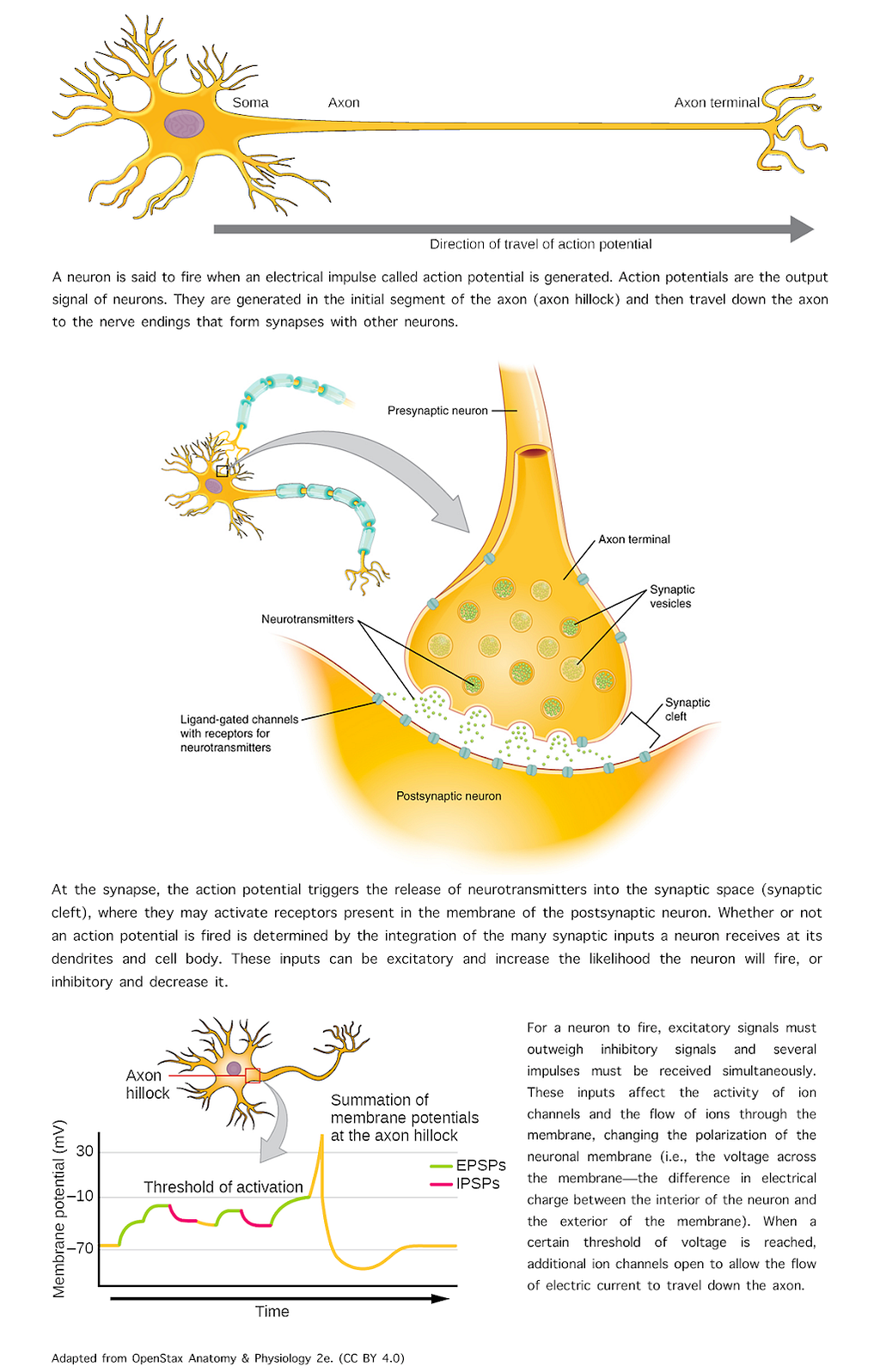
Figure 2 — Action potential. Image adapted from OpenStax, Anatomy & Physiology 2e, Ch.12 The Nervous System and Nervous Tissue (CC BY 4.0)
Whether or not an action potential is fired is determined by the summation of the many synaptic inputs a neuron is receiving and their effect on membrane polarization. In turn, changes in membrane polarization are determined by the properties, distribution, and responsiveness of ion channels in the plasma membrane of the dendrites and soma.
Ion channel activity is the key element of another type of functional plasticity known as intrinsic plasticity. This refers to changes in the excitability of neurons, i.e., the likelihood that a neuron will fire or, in other words, how prone a neuron is to generate an action potential in response to an input signal. Changes in ion channel activity can either facilitate or inhibit neuronal firing [6,7], altering the responsiveness of the neuron to synaptic inputs (i.e., its excitability). The many synaptic inputs a neuron receives affect ion channels differently, but the integration of those many inputs can either tip the balance towards increasing or decreasing neuronal excitability.
Whereas synaptic plasticity changes the strength of a specific synapse and its individual contribution to the overall likelihood that a neuron will fire, intrinsic plasticity changes how the many synaptic inputs a neuron receives are integrated into the overall activity of the neuron. Intrinsic plasticity can thus act as a signal amplifier by increasing the response amplitude and lowering the firing threshold of the neuron, or as a signal attenuator by decreasing the response amplitude and increasing the firing threshold [8].
Importantly, intrinsic excitability, like synaptic strength, can change in an activity-dependent manner as a result of our life experiences. As synaptic function is intrinsically linked to neuronal firing and the electrical properties of dendrites are intrinsically linked to synaptic input, the mechanism of synaptic and intrinsic plasticity influence each other [8]. Intrinsic plasticity is believed to complement and be synergistic with synaptic plasticity in the process of storing information [6,7].
Mechanisms of Structural Plasticity
Mechanisms of structural plasticity range from subtler changes such as synaptic remodeling and myelination of axons, to more dramatic changes such as the generation of new neurons (neurogenesis). The occurrence, extent, and types of structural plasticity varies significantly among different mammalian species and throughout their lifetime; it is unclear whether or not some of these occur in adult humans, as we’ll see.
Synapse Remodeling
In addition to being functionally plastic, synapses are also structurally modifiable through changes in synaptic structures known as dendritic spines [9–11]. These are small protrusions found along the dendrites of many types of neurons (though not all types) (Fig.3) [12].
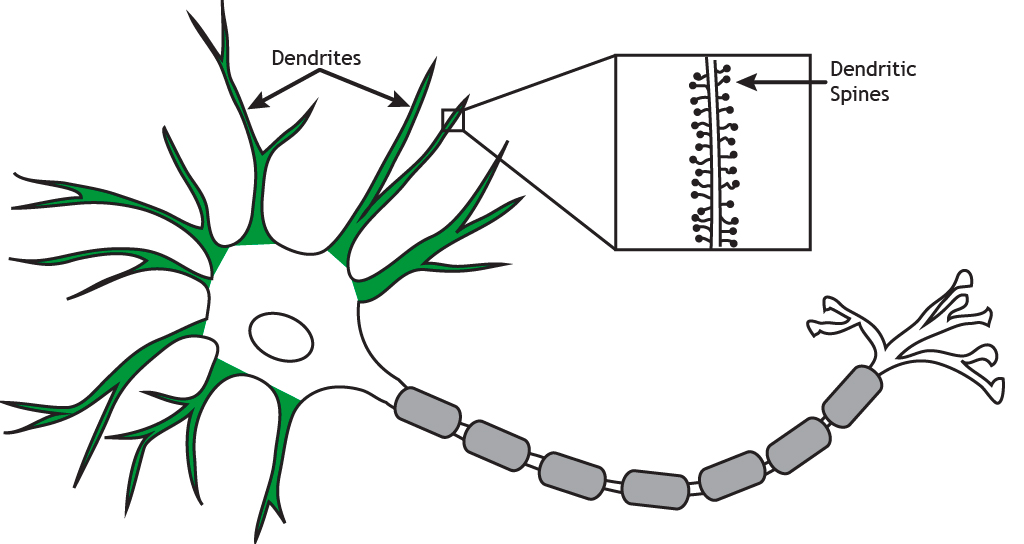
Figure 3 — Dendritic spines. Source: Foundations of Neuroscience by Casey Henley, Ch.1 The Neuron. Michigan State University Open Libraries. (CC BY 4.0)
Spines are formed at a high rate after birth, undergo extensive pruning during postnatal development, and remain largely stable through adulthood, providing a structural substrate for long-term information storage [12–15]. But evidence from animal studies indicates that, even though synaptic remodeling decreases substantially in the adult brain, the capacity for synaptic remodeling remains large enough to have functional significance and lifelong impact on synaptic networks and behavioral adaptation [16–22]. Dendritic spines may swell or shrink, new spines may be formed, spines may be stabilized or pruned [12–15].
Synaptic remodeling often happens in association with functional synaptic plasticity, but on larger timescales [23,24], acting as a structural correlate to functional plasticity. Synaptic strengthening can be accompanied by long-term structural adjustments, including the generation of new dendritic spine next to strengthened synapses [11,21,25], while synaptic weakening can lead to opposite adaptations, such as shrinkage or pruning of dendritic spines [11].
In animal studies, motor task learning and novel sensory experiences triggered a rapid formation of new spines in the adult brain, but most of them ended up being pruned. However, a fraction of those new spines stabilized with repeated stimulation and were integrated into a functional circuit [13,16,17,26]. In the motor learning experiments, the surviving fraction stabilized as motor performance improved, with the level of performance correlating with the amount of retained new spines. Remodeled spines were maintained even after long periods of time without training, indicating that the stabilization of new spines contributed to memory consolidation by creating a structural imprint of the newly learned motor skills [13,26].
This does not mean that the number of spines actually changes with any new experience. In the motor learning experiments, in parallel to new spine formation, the pruning of old spines also took place. This means that the net result of synaptic remodeling processes may actually be the maintenance of total spine density levels when spine formation and spine loss cancel each other out [13,26]. But when spine formation and elimination are differentially affected, spine density may actually change [27]. For example, in another animal study, environmental enrichment (i.e., making the environment more stimulating cognitively and physically) lead to an actual increase in the number of spines [28].
Through synaptic remodeling processes, life experiences may induce small but permanent changes that complement functional plasticity and that refine neuronal circuits to create a stable structural basis for lifelong memory storage and behavioral adaptation [13,15,26].
White Matter and Myelin Plasticity
The CNS is arranged into gray matter and white matter. Gray matter consists primarily (but not only) of neuronal cell bodies, dendrites, and unmyelinated axons, whereas white matter is mostly made up of bundles of myelinated axons that connect different brain regions and form functional circuits. Its white color comes from myelin, the fatty coating that surrounds myelinated axons (Figure 4).
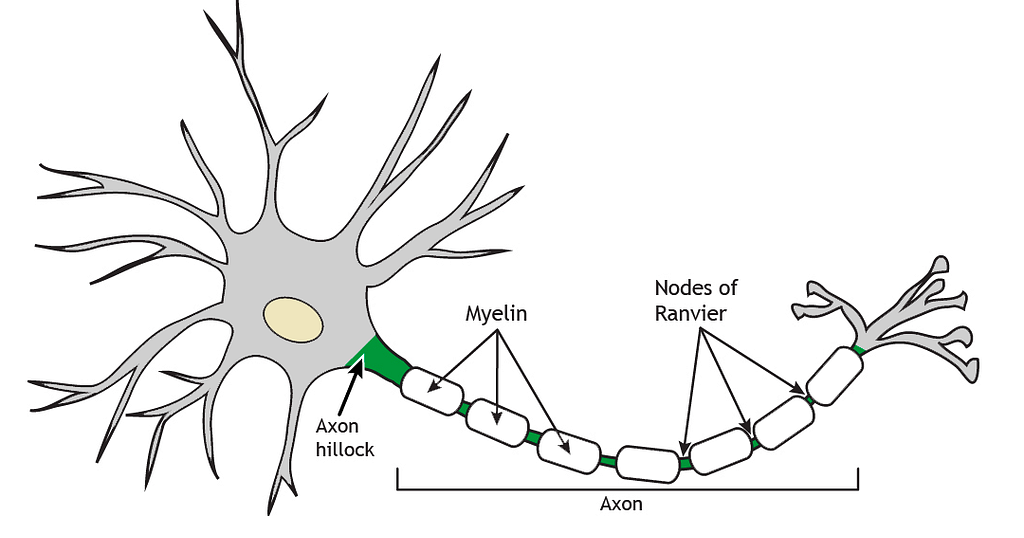
Figure 4 — Myelin. Adapted from: Foundations of Neuroscience by Casey Henley, Ch.1 The Neuron. Michigan State University Open Libraries. (CC BY 4.0)
White matter keeps develops well into adulthood (~30y) and its microstructure starts to decline around middle age (~50y) [29–32], but myelin production by oligodendrocytes (a type of glial cell) and dynamic adjustments in myelination seem to be retained through life [33–36].
Myelin provides electrical insulation to axons and allows for high-speed impulse conduction. Myelin thickness influences conduction speed, which means that changes in myelin production can influence the timing and synchrony of neuronal signals, along with other important aspects of cortical processing. Myelin production is susceptible to activity-dependent changes in neurons, thus allowing for experience-dependent white matter plasticity [37].
Studies in adult mammals have shown that increased neuronal activity can enhance oligodendrocyte generation and promote myelin thickening [38]. It was shown that motor skill learning not only changed white matter microstructure and myelination [39], but also that active myelination was actually necessary for motor skill learning [40,41]. This showed that myelin production is an important element of the neuroplastic changes underlying learning.
Human neuroimaging studies have supported the view that white matter is dynamic also in the human brain as changes in white matter have been linked to a number of learning tasks in adults [42–45]. Through changes in myelin production, white matter plasticity may enable the fine-tuning of neural networks in the adult brain and potentially throughout life, being a mechanism through which life experiences can shape brain function.
Neurogenesis
The generation of new neurons is known as neurogenesis. It is a process that, in the mammalian CNS, is very restricted beyond developmental years and declines substantially with age. Still, neurogenesis has been reported in the adult brain of several mammalian species [46,47]. Whether or not neurogenesis takes place in the human brain is still unclear and generating heated discussion [48,49].
Neurogenesis seems to be possible only in specific brain regions where the molecular and cellular microenvironment allows for neuronal development to occur [50]. These regions are known as neurogenic niches and they harbor pools of neural stem cells (NSCs)-self-renewing, multipotent stem cells of the nervous system from which neurons (and glial cells) can be born.
In adult rodents, there are two canonical neurogenic regions where NSCs reside and neurogenesis is relatively common: (1) a region called subventricular zone (SVZ) from where new neurons migrate to the olfactory bulb, and (2) the subgranular zone (SGZ) of the dentate gyrus (DG) in the hippocampus [51,52]. Neuronal precursors originating from these sites can differentiate into different types of neurons and functionally integrate into local circuits [53]. Neurogenesis outside these two regions is considered to be generally restricted, although evidence of neurogenesis has been found in other regions (with regions varying between studied species), including the neocortex, striatum, amygdala, hypothalamus, substantia nigra, cerebellum and brainstem [54].
The neurogenic capacity of those niches drops exponentially throughout life due to stem cell depletion [55]. This reduction is greater and faster in long-living mammals with larger brains; in sheep and dolphins, for example, it is virtually absent [46,56].
Similarly, in humans, neurogenesis declines dramatically with age. While evidence of hippocampal neurogenesis in the adult human brain has been reported [57,58], other studies have failed to replicate those findings and concluded that, if neurogenesis continues in the adult hippocampus, it must be extremely rare [59,60].
Studying neurogenesis in the human brain is technically and ethically difficult. So, although a lot has been learned about neurogenesis in rodents [61], and despite the hype that findings supporting adult human neurogenesis always generate, the reality is that it’s still unclear whether or to what extent the human brain shares the same capacity [62–64].
Immature Neurons in the Adult Brain
Recently, another type of structural plasticity has been presented as an alternative to the birth of new neurons from NSCs in the adult brain. The hypothesis that has been put forward is that the human adult brain may retain a pool of quiescent immature neurons generated during developmental years but that remain in a persistent state of immaturity. These could potentially be a source of new functional neurons throughout life [65].
In contrast to adult neurogenesis, which decreases in mammals with larger brains and longer lives (e.g., humans, dolphins, and sheep) [56], immature neurons seem to be present in higher numbers in those same mammals. In mammals with smaller brains, non-newly generated immature neurons (i.e., generated during embryonic development and not from stem cells in the adult brain) have been found only in the paleocortex, the evolutionarily oldest part of the cerebral cortex [66].
But in large-brain mammals (including aged non-human primates), non-newly generated immature neurons have been consistently found in the evolutionarily younger neocortex, involved in processing higher cognitive functions, and in brain regions involved in the coupling of emotions and conscious perception, such as the amygdala [67–71]. Evidence has been found for a slow, delayed maturation of those neurons and for their functional integration into pre-existing neural circuits [72–75].
In human adults, markers of immature neurons have been found in the amygdala and hippocampus. Therefore, it is possible that neurons that remain immature for decades may form a substrate for persistent plasticity in the human brain by serving as a reservoir of young neurons that may potentially be integrated into neuronal networks [68,76–78]. This hypothesis is fascinating, but the fact is that it’s still unclear if and to what extent maturation of these neurons does take place in the human brain.
It is possible that this “non-neurogenic” or “suspended neurogenesis” form of supplying new mature neurons in adulthood might have been an evolutionary alternative to neurogenesis in large-brained, long-living mammals. A possible reason is that adult neurogenesis is an “expensive” biological process that is less compatible with the characteristics of larger brains with more complex cognitive functions. This reserve of “stand by” neurons could thus be an alternative pool of new functional neurons that may be integrated throughout life in neural circuits, allowing the adult brain to better cope with cognitive demands or to offset progressive structural decline linked to aging [68,78,79].
Glial Cells as Modulators of Plasticity
Glial cells, which include microglia, astrocytes, and oligodendrocytes, have important functions in the human brain, providing functional and structural support and protection for neurons [80].
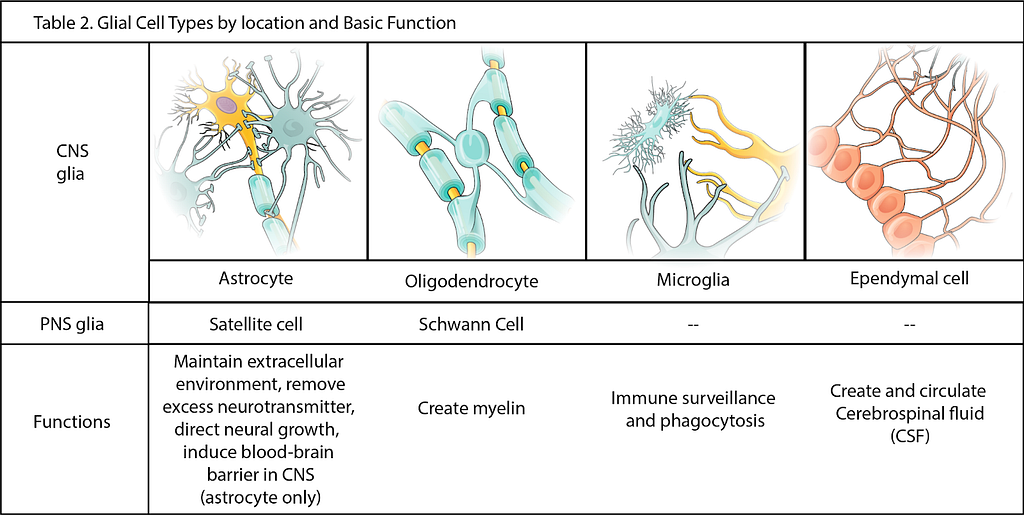
Figure 3 — Glial cells and their main functions. Image adapted from Table 12.2 in Anatomy and Physiology, Chapter 12.2 Nervous Tissue. Oregon State University. (CC BY 4.0)
During development, glial cells play important roles in assisting in the assembly of the nervous system. Once neuronal circuits are formed, glia regulate several aspects of nervous system function throughout life, including neurotransmitter synthesis, neuronal metabolism, immune surveillance, synaptic communication, neuronal network function, or blood brain barrier integrity, for example.
Glial cells are themselves highly plastic-they can proliferate, swell, and grow their cellular extensions, for example. But they also play important parts in neural plasticity. Glial cells are involved in functional synaptic plasticity, in the generation and pruning of synapses, in myelination, and in neuronal excitability, thus influencing the homeostasis and function of the CNS throughout life [81]. Also, morphological modifications of glial cells and changes in glial cell numbers may to some extent underlie structural plastic changes that manifest as increases in gray matter and white matter volume observed in cognitively stimulating contexts, for example [82]. Their coordinated actions provide critical assistance to the plastic adaptability of neurons and neural networks.
Can We Hack Neuroplasticity?
We’ve seen how the brain’s plastic capacity operates at different levels, from molecules to circuits. Neuroplastic changes take place throughout the lifespan, albeit with significant variability in plastic capacity at different ages. Importantly, neuroplasticity is responsive to physiological and environmental factors. Does this mean we can tap into the plastic potential of the brain to improve our cognitive capacity? We try to find some answers in the accompanying article Rewire Your Brain: Science-Backed Methods to Boost Neuroplasticity.
References
1] M. Frotscher, Specificity of interneuronal connections, Ann. Anat. 174 (1992) 377–382. [2] W.C. Abraham, O.D. Jones, D.L. Glanzman, Is plasticity of synapses the mechanism of long-term memory storage?, NPJ Sci Learn. 4 (2019) 9. [3] F. Sengpiel, The critical period, Curr. Biol. 17 (2007) R742–3. [4] B. Hassel, R. Dingledine, Chapter 17 — Glutamate and Glutamate Receptors, in: S.T. Brady, G.J. Siegel, R.W. Albers, D.L. Price (Eds.), Basic Neurochemistry (Eighth Edition), Academic Press, New York, 2012: pp. 342–366. [5] J.Z. Tsien, Chapter 56 — Learning and Memory, in: S.T. Brady, G.J. Siegel, R.W. Albers, D.L. Price (Eds.), Basic Neurochemistry (Eighth Edition), Academic Press, New York, 2012: pp. 963–981. [6] C.R. Gallistel, L.D. Matzel, The neuroscience of learning: beyond the Hebbian synapse, Annu. Rev. Psychol. 64 (2013) 169–200. [7] H.G. Shim, Y.-S. Lee, S.J. Kim, The Emerging Concept of Intrinsic Plasticity: Activity-dependent Modulation of Intrinsic Excitability in Cerebellar Purkinje Cells and Motor Learning, Exp. Neurobiol. 27 (2018) 139–154. [8] H.K. Titley, N. Brunel, C. Hansel, Toward a Neurocentric View of Learning, Neuron. 95 (2017) 19–32. [9] C.R. Bramham, Local protein synthesis, actin dynamics, and LTP consolidation, Curr. Opin. Neurobiol. 18 (2008) 524–531. [10] T. Tada, M. Sheng, Molecular mechanisms of dendritic spine morphogenesis, Curr. Opin. Neurobiol. 16 (2006) 95–101. [11] Q. Zhou, K.J. Homma, M.-M. Poo, Shrinkage of dendritic spines associated with long-term depression of hippocampal synapses, Neuron. 44 (2004) 749–757. [12] P. Caroni, F. Donato, D. Muller, Structural plasticity upon learning: regulation and functions, Nat. Rev. Neurosci. 13 (2012) 478–490. [13] G. Yang, F. Pan, W.-B. Gan, Stably maintained dendritic spines are associated with lifelong memories, Nature. 462 (2009) 920–924. [14] J. Bourne, K.M. Harris, Do thin spines learn to be mushroom spines that remember?, Curr. Opin. Neurobiol. 17 (2007) 381–386. [15] K. Runge, C. Cardoso, A. de Chevigny, Dendritic Spine Plasticity: Function and Mechanisms, Front. Synaptic Neurosci. 12 (2020) 36. [16] A.J.G.D. Holtmaat, J.T. Trachtenberg, L. Wilbrecht, G.M. Shepherd, X. Zhang, G.W. Knott, K. Svoboda, Transient and persistent dendritic spines in the neocortex in vivo, Neuron. 45 (2005) 279–291. [17] J.T. Trachtenberg, B.E. Chen, G.W. Knott, G. Feng, J.R. Sanes, E. Welker, K. Svoboda, Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex, Nature. 420 (2002) 788–794. [18] J. Grutzendler, N. Kasthuri, W.-B. Gan, Long-term dendritic spine stability in the adult cortex, Nature. 420 (2002) 812–816. [19] A. Holtmaat, K. Svoboda, Experience-dependent structural synaptic plasticity in the mammalian brain, Nat. Rev. Neurosci. 10 (2009) 647–658. [20] M. Matsuzaki, N. Honkura, G.C.R. Ellis-Davies, H. Kasai, Structural basis of long-term potentiation in single dendritic spines, Nature. 429 (2004) 761–766. [21] R. Yuste, T. Bonhoeffer, Morphological changes in dendritic spines associated with long-term synaptic plasticity, Annu. Rev. Neurosci. 24 (2001) 1071–1089. [22] M. Bosch, Y. Hayashi, Structural plasticity of dendritic spines, Curr. Opin. Neurobiol. 22 (2012) 383–388. [23] D.B. Chklovskii, B.W. Mel, K. Svoboda, Cortical rewiring and information storage, Nature. 431 (2004) 782–788. [24] M. Butz, F. Wörgötter, A. van Ooyen, Activity-dependent structural plasticity, Brain Res. Rev. 60 (2009) 287–305. [25] N. Toni, P.A. Buchs, I. Nikonenko, C.R. Bron, D. Muller, LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite, Nature. 402 (1999) 421–425. [26] T. Xu, X. Yu, A.J. Perlik, W.F. Tobin, J.A. Zweig, K. Tennant, T. Jones, Y. Zuo, Rapid formation and selective stabilization of synapses for enduring motor memories, Nature. 462 (2009) 915–919. [27] S.B. Hofer, T.D. Mrsic-Flogel, T. Bonhoeffer, M. Hübener, Experience leaves a lasting structural trace in cortical circuits, Nature. 457 (2009) 313–317. [28] E. Bednarek, P. Caroni, β-Adducin is required for stable assembly of new synapses and improved memory upon environmental enrichment, Neuron. 69 (2011) 1132–1146. [29] C. Lebel, C. Beaulieu, Longitudinal development of human brain wiring continues from childhood into adulthood, J. Neurosci. 31 (2011) 10937–10947. [30] E.R. Sowell, P.M. Thompson, C.J. Holmes, T.L. Jernigan, A.W. Toga, In vivo evidence for post-adolescent brain maturation in frontal and striatal regions, Nat. Neurosci. 2 (1999) 859–861. [31] C.E. Sexton, K.B. Walhovd, A.B. Storsve, C.K. Tamnes, L.T. Westlye, H. Johansen-Berg, A.M. Fjell, Accelerated changes in white matter microstructure during aging: a longitudinal diffusion tensor imaging study, J. Neurosci. 34 (2014) 15425–15436. [32] C. Lebel, M. Gee, R. Camicioli, M. Wieler, W. Martin, C. Beaulieu, Diffusion tensor imaging of white matter tract evolution over the lifespan, Neuroimage. 60 (2012) 340–352. [33] M.S.Y. Yeung, S. Zdunek, O. Bergmann, S. Bernard, M. Salehpour, K. Alkass, S. Perl, J. Tisdale, G. Possnert, L. Brundin, H. Druid, J. Frisén, Dynamics of oligodendrocyte generation and myelination in the human brain, Cell. 159 (2014) 766–774. [34] K.M. Young, K. Psachoulia, R.B. Tripathi, S.-J. Dunn, L. Cossell, D. Attwell, K. Tohyama, W.D. Richardson, Oligodendrocyte dynamics in the healthy adult CNS: evidence for myelin remodeling, Neuron. 77 (2013) 873–885. [35] R.A. Hill, A.M. Li, J. Grutzendler, Lifelong cortical myelin plasticity and age-related degeneration in the live mammalian brain, Nat. Neurosci. 21 (2018) 683–695. [36] E.G. Hughes, J.L. Orthmann-Murphy, A.J. Langseth, D.E. Bergles, Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex, Nat. Neurosci. 21 (2018) 696–706. [37] R.D. Fields, A new mechanism of nervous system plasticity: activity-dependent myelination, Nat. Rev. Neurosci. 16 (2015) 756–767. [38] E.M. Gibson, D. Purger, C.W. Mount, A.K. Goldstein, G.L. Lin, L.S. Wood, I. Inema, S.E. Miller, G. Bieri, J.B. Zuchero, B.A. Barres, P.J. Woo, H. Vogel, M. Monje, Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain, Science. 344 (2014) 1252304. [39] C. Sampaio-Baptista, A.A. Khrapitchev, S. Foxley, T. Schlagheck, J. Scholz, S. Jbabdi, G.C. DeLuca, K.L. Miller, A. Taylor, N. Thomas, J. Kleim, N.R. Sibson, D. Bannerman, H. Johansen-Berg, Motor skill learning induces changes in white matter microstructure and myelination, J. Neurosci. 33 (2013) 19499–19503. [40] I.A. McKenzie, D. Ohayon, H. Li, J.P. de Faria, B. Emery, K. Tohyama, W.D. Richardson, Motor skill learning requires active central myelination, Science. 346 (2014) 318–322. [41] L. Xiao, D. Ohayon, I.A. McKenzie, A. Sinclair-Wilson, J.L. Wright, A.D. Fudge, B. Emery, H. Li, W.D. Richardson, Rapid production of new oligodendrocytes is required in the earliest stages of motor-skill learning, Nat. Neurosci. 19 (2016) 1210–1217. [42] J. Scholz, M.C. Klein, T.E.J. Behrens, H. Johansen-Berg, Training induces changes in white-matter architecture, Nat. Neurosci. 12 (2009) 1370–1371. [43] S.L. Bengtsson, Z. Nagy, S. Skare, L. Forsman, H. Forssberg, F. Ullén, Extensive piano practicing has regionally specific effects on white matter development, Nat. Neurosci. 8 (2005) 1148–1150. [44] A.A. Schlegel, J.J. Rudelson, P.U. Tse, White matter structure changes as adults learn a second language, J. Cogn. Neurosci. 24 (2012) 1664–1670. [45] M. Carreiras, M.L. Seghier, S. Baquero, A. Estévez, A. Lozano, J.T. Devlin, C.J. Price, An anatomical signature for literacy, Nature. 461 (2009) 983–986. [46] R. Parolisi, B. Cozzi, L. Bonfanti, Humans and Dolphins: Decline and Fall of Adult Neurogenesis, Front. Neurosci. 12 (2018) 497. [47] L. Bonfanti, I. Amrein, Editorial: Adult Neurogenesis: Beyond Rats and Mice, Front. Neurosci. 12 (2018) 904. [48] E.P. Moreno-Jiménez, J. Terreros-Roncal, M. Flor-García, A. Rábano, M. Llorens-Martín, Evidences for Adult Hippocampal Neurogenesis in Humans, J. Neurosci. 41 (2021) 2541–2553. [49] S.F. Sorrells, M.F. Paredes, Z. Zhang, G. Kang, O. Pastor-Alonso, S. Biagiotti, C.E. Page, K. Sandoval, A. Knox, A. Connolly, E.J. Huang, J.M. Garcia-Verdugo, M.C. Oldham, Z. Yang, A. Alvarez-Buylla, Positive Controls in Adults and Children Support That Very Few, If Any, New Neurons Are Born in the Adult Human Hippocampus, J. Neurosci. 41 (2021) 2554–2565. [50] T.D. Palmer, A.R. Willhoite, F.H. Gage, Vascular niche for adult hippocampal neurogenesis, J. Comp. Neurol. 425 (2000) 479–494. [51] F.H. Gage, Mammalian Neural Stem Cells, Science. 287 (2000) 1433–1438. [52] D.A. Lim, A. Alvarez-Buylla, The Adult Ventricular-Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis, Cold Spring Harb. Perspect. Biol. 8 (2016). [53] J.B. Aimone, Y. Li, S.W. Lee, G.D. Clemenson, W. Deng, F.H. Gage, Regulation and function of adult neurogenesis: from genes to cognition, Physiol. Rev. 94 (2014) 991–1026. [54] D.M. Feliciano, A. Bordey, L. Bonfanti, Noncanonical Sites of Adult Neurogenesis in the Mammalian Brain, Cold Spring Harb. Perspect. Biol. 7 (2015). [55] K. Smith, M.V. Semënov, The impact of age on number and distribution of proliferating cells in subgranular zone in adult mouse brain, IBRO Rep. 6 (2019) 18–30. [56] M.F. Paredes, S.F. Sorrells, J.M. Garcia-Verdugo, A. Alvarez-Buylla, Brain size and limits to adult neurogenesis, J. Comp. Neurol. 524 (2016) 646–664. [57] M. Boldrini, C.A. Fulmore, A.N. Tartt, L.R. Simeon, I. Pavlova, V. Poposka, G.B. Rosoklija, A. Stankov, V. Arango, A.J. Dwork, R. Hen, J.J. Mann, Human Hippocampal Neurogenesis Persists throughout Aging, Cell Stem Cell. 22 (2018) 589–599.e5. [58] E.P. Moreno-Jiménez, M. Flor-García, J. Terreros-Roncal, A. Rábano, F. Cafini, N. Pallas-Bazarra, J. Ávila, M. Llorens-Martín, Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer’s disease, Nat. Med. 25 (2019) 554–560. [59] S.F. Sorrells, M.F. Paredes, A. Cebrian-Silla, K. Sandoval, D. Qi, K.W. Kelley, D. James, S. Mayer, J. Chang, K.I. Auguste, E.F. Chang, A.J. Gutierrez, A.R. Kriegstein, G.W. Mathern, M.C. Oldham, E.J. Huang, J.M. Garcia-Verdugo, Z. Yang, A. Alvarez-Buylla, Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults, Nature. 555 (2018) 377–381. [60] S. Cipriani, I. Ferrer, E. Aronica, G.G. Kovacs, C. Verney, J. Nardelli, S. Khung, A.-L. Delezoide, I. Milenkovic, S. Rasika, P. Manivet, J.-L. Benifla, N. Deriot, P. Gressens, H. Adle-Biassette, Hippocampal Radial Glial Subtypes and Their Neurogenic Potential in Human Fetuses and Healthy and Alzheimer’s Disease Adults, Cereb. Cortex. 28 (2018) 2458–2478. [61] A.M. Bond, G.-L. Ming, H. Song, Adult Mammalian Neural Stem Cells and Neurogenesis: Five Decades Later, Cell Stem Cell. 17 (2015) 385–395. [62] A. Duque, R. Spector, A balanced evaluation of the evidence for adult neurogenesis in humans: implication for neuropsychiatric disorders, Brain Struct. Funct. 224 (2019) 2281–2295. [63] D. Petrik, J.M. Encinas, Perspective: Of Mice and Men — How Widespread Is Adult Neurogenesis?, Front. Neurosci. 13 (2019) 923. [64] J.S. Snyder, Recalibrating the Relevance of Adult Neurogenesis, Trends Neurosci. 42 (2019) 164–178. [65] C. La Rosa, R. Parolisi, L. Bonfanti, Brain Structural Plasticity: From Adult Neurogenesis to Immature Neurons, Front. Neurosci. 14 (2020) 75. [66] M.A. Gómez-Climent, E. Castillo-Gómez, E. Varea, R. Guirado, J.M. Blasco-Ibáñez, C. Crespo, F.J. Martínez-Guijarro, J. Nácher, A population of prenatally generated cells in the rat paleocortex maintains an immature neuronal phenotype into adulthood, Cereb. Cortex. 18 (2008) 2229–2240. [67] C. La Rosa, F. Cavallo, A. Pecora, M. Chincarini, U. Ala, C.G. Faulkes, J. Nacher, B. Cozzi, C.C. Sherwood, I. Amrein, L. Bonfanti, Phylogenetic variation in cortical layer II immature neuron reservoir of mammals, Elife. 9 (2020). [68] O. Palazzo, C. La Rosa, M. Piumatti, L. Bonfanti, Do large brains of long-living mammals prefer non-newly generated, immature neurons?, Neural Regeneration Res. 13 (2018) 633–634. [69] M. Piumatti, O. Palazzo, C. La Rosa, P. Crociara, R. Parolisi, F. Luzzati, F. Lévy, L. Bonfanti, Non-Newly Generated, “Immature” Neurons in the Sheep Brain Are Not Restricted to Cerebral Cortex, The Journal of Neuroscience. 38 (2018) 826–842. [70] X.-M. Zhang, Y. Cai, Y. Chu, E.-Y. Chen, J.-C. Feng, X.-G. Luo, K. Xiong, R.G. Struble, R.W. Clough, P.R. Patrylo, J.H. Kordower, X.-X. Yan, Doublecortin-expressing cells persist in the associative cerebral cortex and amygdala in aged nonhuman primates, Front. Neuroanat. 3 (2009) 17. [71] J. Bloch, M. Kaeser, Y. Sadeghi, E.M. Rouiller, D.E. Redmond Jr, J.-F. Brunet, Doublecortin-positive cells in the adult primate cerebral cortex and possible role in brain plasticity and development, J. Comp. Neurol. 519 (2011) 775–789. [72] S.J. Kohler, N.I. Williams, G.B. Stanton, J.L. Cameron, W.T. Greenough, Maturation time of new granule cells in the dentate gyrus of adult macaque monkeys exceeds six months, Proc. Natl. Acad. Sci. U. S. A. 108 (2011) 10326–10331. [73] M. Brus, M. Meurisse, G. Gheusi, M. Keller, P.M. Lledo, F. Lévy, Dynamics of olfactory and hippocampal neurogenesis in adult sheep, J. Comp. Neurol. 521 (2013) 169–188. [74] P. Rotheneichner, M. Belles, B. Benedetti, R. König, D. Dannehl, C. Kreutzer, P. Zaunmair, M. Engelhardt, L. Aigner, J. Nacher, S. Couillard-Despres, Cellular Plasticity in the Adult Murine Piriform Cortex: Continuous Maturation of Dormant Precursors Into Excitatory Neurons, Cereb. Cortex. 28 (2018) 2610–2621. [75] B. Benedetti, D. Dannehl, R. König, S. Coviello, C. Kreutzer, P. Zaunmair, D. Jakubecova, T.M. Weiger, L. Aigner, J. Nacher, M. Engelhardt, S. Couillard-Després, Functional Integration of Neuronal Precursors in the Adult Murine Piriform Cortex, Cereb. Cortex. 30 (2020) 1499–1515. [76] U. Martí-Mengual, E. Varea, C. Crespo, J.M. Blasco-Ibáñez, J. Nacher, Cells expressing markers of immature neurons in the amygdala of adult humans, Eur. J. Neurosci. 37 (2013) 10–22. [77] T. Seki, T. Hori, H. Miyata, M. Maehara, T. Namba, Analysis of proliferating neuronal progenitors and immature neurons in the human hippocampus surgically removed from control and epileptic patients, Sci. Rep. 9 (2019) 18194. [78] C. La Rosa, M. Ghibaudi, L. Bonfanti, Newly Generated and Non-Newly Generated “Immature” Neurons in the Mammalian Brain: A Possible Reservoir of Young Cells to Prevent Brain Aging and Disease?, J. Clin. Med. Res. 8 (2019). [79] L. Bonfanti, C.J. Charvet, Brain Plasticity in Humans and Model Systems: Advances, Challenges, and Future Directions, Int. J. Mol. Sci. 22 (2021). [80] N.J. Allen, D.A. Lyons, Glia as architects of central nervous system formation and function, Science. 362 (2018) 181–185. [81] L. Sancho, M. Contreras, N.J. Allen, Glia as sculptors of synaptic plasticity, Neurosci. Res. 167 (2021) 17–29. https://doi.org/10.1016/s0940-9602(11)80254-2.
https://doi.org/10.1038/s41539-019-0048-y.
https://doi.org/10.1016/j.cub.2007.06.017.
https://doi.org/10.1016/B978-0-12-374947-5.00017-1.
https://doi.org/10.1016/B978-0-12-374947-5.00056-0.
https://doi.org/10.1146/annurev-psych-113011-143807.
https://doi.org/10.5607/en.2018.27.3.139.
https://doi.org/10.1016/j.neuron.2017.05.021.
https://doi.org/10.1016/j.conb.2008.09.013.
https://doi.org/10.1016/j.conb.2005.12.001.
https://doi.org/10.1016/j.neuron.2004.11.011.
https://doi.org/10.1038/nrn3258.
https://doi.org/10.1038/nature08577.
https://doi.org/10.1016/j.conb.2007.04.009.
https://doi.org/10.3389/fnsyn.2020.00036.
https://doi.org/10.1016/j.neuron.2005.01.003.
https://doi.org/10.1038/nature01273.
https://doi.org/10.1038/nature01276.
https://doi.org/10.1038/nrn2699.
https://doi.org/10.1038/nature02617.
https://doi.org/10.1146/annurev.neuro.24.1.1071.
https://doi.org/10.1016/j.conb.2011.09.002.
https://doi.org/10.1038/nature03012.
https://doi.org/10.1016/j.brainresrev.2008.12.023.
https://doi.org/10.1038/46574.
https://doi.org/10.1038/nature08389.
https://doi.org/10.1038/nature07487.
https://doi.org/10.1016/j.neuron.2011.02.034.
https://doi.org/10.1523/JNEUROSCI.5302-10.2011.
https://doi.org/10.1038/13154.
https://doi.org/10.1523/JNEUROSCI.0203-14.2014.
https://doi.org/10.1016/j.neuroimage.2011.11.094.
https://doi.org/10.1016/j.cell.2014.10.011.
https://doi.org/10.1016/j.neuron.2013.01.006.
https://doi.org/10.1038/s41593-018-0120-6.
https://doi.org/10.1038/s41593-018-0121-5.
https://doi.org/10.1038/nrn4023.
https://doi.org/10.1126/science.1252304.
https://doi.org/10.1523/JNEUROSCI.3048-13.2013. https://doi.org/10.1126/science.1254960.
https://doi.org/10.1038/nn.4351.
https://doi.org/10.1038/nn.2412.
https://doi.org/10.1038/nn1516.
https://doi.org/10.1162/jocn_a_00240.
https://doi.org/10.1038/nature08461.
https://doi.org/10.3389/fnins.2018.00497.
https://doi.org/10.3389/fnins.2018.00904.
https://doi.org/10.1523/JNEUROSCI.0675-20.2020.
https://doi.org/10.1523/JNEUROSCI.0676-20.2020.
https://onlinelibrary.wiley.com/doi/abs/10.1002/1096-9861(20001002)425:4%3C479::AID-CNE2%3E3.0.CO;2-3.
https://doi.org/10.1126/science.287.5457.1433.
https://doi.org/10.1101/cshperspect.a018820.
https://doi.org/10.1152/physrev.00004.2014.
https://doi.org/10.1101/cshperspect.a018846.
https://doi.org/10.1016/j.ibror.2018.12.002.
https://doi.org/10.1002/cne.23896.
https://doi.org/10.1016/j.stem.2018.03.015.
https://doi.org/10.1038/s41591-019-0375-9.
https://doi.org/10.1038/nature25975.
https://doi.org/10.1093/cercor/bhy096.
https://doi.org/10.1016/j.stem.2015.09.003.
https://doi.org/10.1007/s00429-019-01917-6.
https://doi.org/10.3389/fnins.2019.00923.
https://doi.org/10.1016/j.tins.2018.12.001.
https://doi.org/10.3389/fnins.2020.00075.
https://doi.org/10.1093/cercor/bhm255.
https://doi.org/10.7554/eLife.55456.
https://doi.org/10.4103/1673-5374.230282.
https://doi.org/10.1523/jneurosci.1781-17.2017.
https://doi.org/10.3389/neuro.05.017.2009.
https://doi.org/10.1002/cne.22547.
https://doi.org/10.1073/pnas.1017099108.
https://doi.org/10.1002/cne.23169.
https://doi.org/10.1093/cercor/bhy087.
https://doi.org/10.1093/cercor/bhz181.
https://doi.org/10.1111/ejn.12016.
https://doi.org/10.1038/s41598-019-54684-z.
https://doi.org/10.3390/jcm8050685.
https://doi.org/10.3390/ijms22179358.
https://doi.org/10.1126/science.aat0473.
https://doi.org/10.1016/j.neures.2020.11.005.
[82] R.J. Zatorre, R.D. Fields, H. Johansen-Berg, Plasticity in gray and white: neuroimaging changes in brain structure during learning, Nat. Neurosci. 15 (2012) 528–536. https://doi.org/10.1038/nn.3045.
Originally published at https://neurohacker.com.
Tagged with :